Le Monde des Mitochondries |
Des Granules d'ARN dans la Mitochondrie |
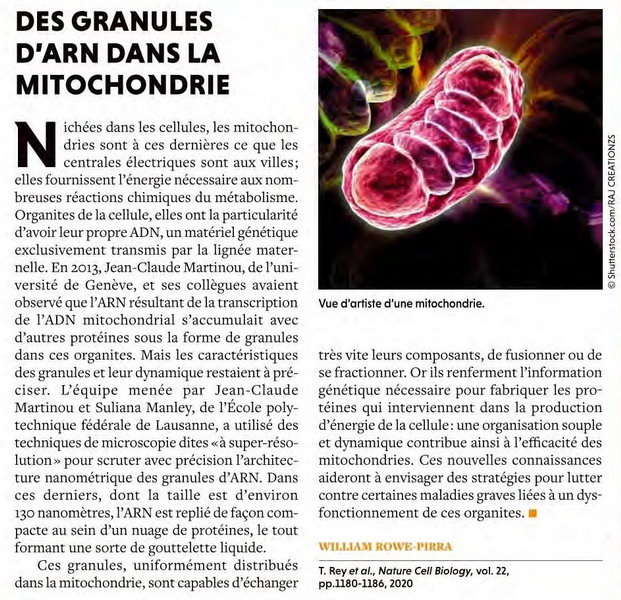
W.R.-P. - POUR LA SCIENCE N°518 > Décembre > 2020 |
Les Mitochondries |
Les mitochondries (du grec mitos, fil et chondros, grain) sont des structures intracellulaires (organites), dont la taille est de l'ordre du micromètre. Leur rôle physiologique est primordial, puisque c'est dans les mitochondries que l'énergie fournie par les molécules organiques est récupérée puis stockée sous forme d'ATP, la source principale d'énergie pour la cellule eucaryote, par le processus de phosphorylation oxydative.
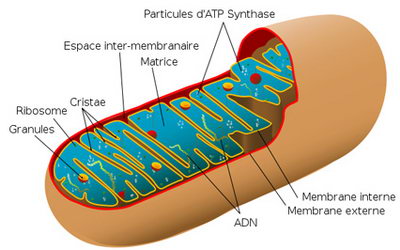
 STRUCTURE : Les mitochondries ont une dimension de 1-2 à 10 µm de long et de 0,5 à 1 µm de large. Elles se composent de 2 membranes mitochondriales, une externe et une interne, qui délimitent trois milieux : le milieu extra-mitochondrial (cytoplasme de la cellule), l'espace inter-membranaire et la matrice. Chacune est de l'ordre des 6 nm et l'espace intermembranaire est de 7 nm. Mitochondries observées en microscopie électronique à transmission (->).
STRUCTURE : Les mitochondries ont une dimension de 1-2 à 10 µm de long et de 0,5 à 1 µm de large. Elles se composent de 2 membranes mitochondriales, une externe et une interne, qui délimitent trois milieux : le milieu extra-mitochondrial (cytoplasme de la cellule), l'espace inter-membranaire et la matrice. Chacune est de l'ordre des 6 nm et l'espace intermembranaire est de 7 nm. Mitochondries observées en microscopie électronique à transmission (->).
- La membrane externe est formée de 60 % de protéines et de 40 % de lipides polaires. Elle contient de nombreuses protéines appelées porines (VDAC) qui forment des canaux aqueux au travers de la membrane. La porine (protéines transmembranaires composé de 16 feuillets béta formant les canaux protéiques traversant la couche bimoléculaire de lipides) laisse passer toutes les molécules hydrophiles d'une masse moléculaire inférieure à 10.000 daltons (anions, cations, les acides gras, le pyruvate, les nucléotides le traversent). La membrane externe présente des complexes TOM constitués de plusieurs sous-unités protéiques dont des récepteurs et des canaux aqueux qui permettent l'entrée des protéines d'origine nucléaire dans la mitochondrie, ou l'insertion de ces mêmes protéines dans la membrane externe.
- La membrane interne est beaucoup moins perméable que la membrane externe. Elle est composée de 75 % de protéines et de 25 % de lipides. Elle contient en quantité un phospholipide double, la cardiolipine, renfermant 4 acides gras rendant cette membrane imperméable aux ions. Les autres molécules doivent passer par un transporteur pour traverser la membrane interne. La membrane interne présente des complexes TIM 23, TIM 22, et OXA. Le TIM 23 permet l'entrée de protéines situées dans l'espace inter-membranaire dans la matrice mitochondriale et dans la membrane interne. Le TIM 22 permet l'insertion des protéines dans la membrane interne et notamment des protéines à plusieurs domaines transmembranaires. Le complexe OXA permet la sortie de la matrice pour certaines protéines d'origine mitochondriale.
La membrane interne forme des invaginations qui apparaissent sous forme de crêtes ou replis au microscope électronique. Ces crêtes augmentent la surface de la membrane et donc de capacité de phosphorylation oxydative. Grâce à cette caractéristique on peut déduire que si une mitochondrie possède beaucoup de crêtes c'est que la cellule a besoin d'une grande quantité d'énergie et donc elle pourra produire plus d'ATP (cellule en activité). On retrouve également à son niveau des protéines de transport spécifiques pour les petites molécules utilisées par la matrice, les enzymes de la chaîne respiratoire, l'ATP-synthase ou complexe F0-F1 visible au microscope électronique sous forme de protubérance interne.
ORIGINE : Une mitochondrie ne peut provenir que de la croissance et de la division d'une autre mitochondrie déjà existante. Normalement, avant la division cellulaire, la mitochondrie double sa masse puis se scinde en deux. Elles sont aussi capables de fusionner entre elles. Cette division débute par l'apparition d'un sillon de division sur la membrane interne. Elle a lieu pendant toute l'interphase et nécessite l'intervention de la protéine DRP1 (voisine de la dynamine). La réplication de l'ADN mitochondrial n'est pas limitée à la phase S du cycle cellulaire. Le nombre de mitochondries par cellule est régulé par l'activité cellulaire. Par exemple, une cellule musculaire au repos contient 5 à 10 fois moins de mitochondries qu'une cellule musculaire activée en permanence.
Le fait que la mitochondrie possède son ADN propre, comme les chloroplastes, indique une origine exogène : il est maintenant admis que les mitochondries proviennent de l'endosymbiose d'une a-protéobactérie il y a environ 2 milliards d'années. La théorie endosymbiotique de l'origine des mitochondrie, a été développée et argumentée par Lynn Margulis dès 1966, puis a été appuyée par la découverte de l'ADN spécifique des mitochondries en 1980. Il semble qu'au cours de l'évolution l'ADN originel de la bactérie ait subi diverses évolutions, perdu un grand nombres de gènes, parfois transféré dans l'ADN de la cellule hôte. Parallèlement à ce report de la synthèse de certaines protéines vers l'hôte, ce dernier a développé un arsenal de translocases, enzymes permettant le transfert de ces protéines vers la matrice mitochondriale.
 LE GÉNOME MITOCHODRIAL : Selon la théorie endosymbiotique, les mitochondries possèderaient une origine monophylétique unique. Une cellule eucaryote primitive (ou une archea) aurait intégré un endosymbionte procaryote il y a environ 1,5 à 2 milliards d'années, lorsque que l'atmosphère primitive s'est enrichie en oxygène. Les études phylogénétiques indiquent que cet endosymbionte est apparenté aux alpha-protéobactéries, le plus proche parent de la mitochondrie connu actuellement étant Rickettsia prowazekii, un parasite intracellulaire obligatoire. Au cours de l'évolution, la majorité des gènes de l'endosymbionte originel auraient été perdus ou bien transférés vers le noyau de la cellule eucaryote hôte. En effet, les nombreux pseudogènes mitochondriaux présents dans le génome attestent d'un processus de transfert tout au long de l'évolution. Vue détaillée d'une mitochondrie (->)
LE GÉNOME MITOCHODRIAL : Selon la théorie endosymbiotique, les mitochondries possèderaient une origine monophylétique unique. Une cellule eucaryote primitive (ou une archea) aurait intégré un endosymbionte procaryote il y a environ 1,5 à 2 milliards d'années, lorsque que l'atmosphère primitive s'est enrichie en oxygène. Les études phylogénétiques indiquent que cet endosymbionte est apparenté aux alpha-protéobactéries, le plus proche parent de la mitochondrie connu actuellement étant Rickettsia prowazekii, un parasite intracellulaire obligatoire. Au cours de l'évolution, la majorité des gènes de l'endosymbionte originel auraient été perdus ou bien transférés vers le noyau de la cellule eucaryote hôte. En effet, les nombreux pseudogènes mitochondriaux présents dans le génome attestent d'un processus de transfert tout au long de l'évolution. Vue détaillée d'une mitochondrie (->)
Le matériel génétique (ADN mitochondrial) de la mitochondrie (qui est la seule partie des cellules animales à posséder son propre ADN, en plus du noyau) sert souvent dans les recherches phylogénétiques. Le génome mitochondrial (ADNmt) humain est circulaire et composé de 16.569 paires de bases, dont 13 cistrons codant des ARNms, 22 gènes pour des ARNts et 2 gènes pour des ARNrs.
Le génome mitochondrial peut être très différent d'une espèce à l'autre, il est extrêmement dynamique, il est souvent hétéroplasmique, c'est-à-dire qu'il coexiste différentes formes au sein de la même mitochondrie. Il peut être trouvé sous forme circulaire ou linéaire, double ou simple brin. Ces différentes formes sont, entre autre, les produits de la réplication du génome mitochondrial par un mécanisme de cercle roulant, mais aussi d'un mécanisme de réplication recombinaison-dépendant, similaire à la réplication du phage T4. Les génomes mitochondriaux sont habituellement représentés sous forme circulaire, le "cercle maître" qui correspond à la molécule décrivant le mieux le génome. Les ribosomes mitochondriaux ou mitoribosomes sont différents des ribosomes de la cellule : ils sont plus petits (70S au lieu de 80S). Le code génétique employé pour la synthèse des protéines peut être différent de celui utilisé dans les synthèses cytosoliques. Chez les vértébrés 4 codons sur 64 ont une signification différente, dont le codon UGA qui est transcrit dans le cytosol en codon stop mais dans la matrice UGA est transcrit en tryptophane (Trp/W), AGG et AGA codent un codon STOP au lieu d'une arginine (Arg/R) et AUA code la méthionine (Met/M) au lieu de l'isoleucine (Ile/I). L'ADN mitochondrial peut aussi se répliquer.
Chez les animaux, lors de la reproduction sexuée, les mitochondries du spermatozoïde pourraient passer dans l'ovocyte, mais le nombre de mitochondries ainsi transférées reste très faible en comparaison de celles déjà présentes dans l'ovocyte. Autrement dit, la quasi totalité des mitochondries de la cellule-ouf provient du gamète femelle. L'étude de l'ADN mitochondrial humain permet donc de retracer les relations généalogiques entre les individus seulement selon la voie maternelle. Certaines études ont ainsi pu décrire un génome mitochondrial ancestral duquel descendraient tous les génomes mitochondriaux de l'humanité. L'individu femelle supposé qui portait ce génome a été dénommé ève mitochondriale. Ce terme biblique reste toutefois trompeur, il est en effet très peu probable que l'humanité ait un unique ancêtre féminin et les récentes études prouvant le transfert de mitochondries provenant des spermatozoïdes lors de la fécondation, remet en cause cette théorie.
LE PROTÉOME MITOCHONDRIAL : La taille du protéome mitochondrial humain est estimée à plus d'un millier de protéines, dont actuellement la moitié est identifiée. Seules 13 protéines sont codées par l'ADN mitochondrial, vestige du génome de l'endosymbionte. Toutes les autres protéines sont codées par le génome nucléaire. Chez la levure 50-60 % des protéines mitochondriales ont des homologues chez les procaryotes alors que 40-50 % n'en ont pas. Les protéines mitochondriales possédant un homologue procaryote résultent probablement du transfert des gènes de l'endosymbionte vers le noyau tandis que les protéines non homologues a des protéines procaryotes résultent d'un phénomène "d'enrichissement" du protéome mitochondrial par de nouvelles protéines et donc de nouvelles fonctions.
Les protéines mitochondriales codées par le génome nucléaire (ou protéines mitochondriales nucléaires) sont importées à l'intérieur de la matrice mitochondriale par différents mécanismes possibles :
- des complexes d'importation (3 sur la membrane interne, 2 sur la membrane externe) ;
- un peptide signal (environ 15 à 30 acides aminés) en position N-terminale de la protéine qui permet sa reconnaissance et son importation dans la mitochondrie,
- grâce à un apport énergétique.
Néanmoins, une part d'environ 10 % est directement synthétisée dans la matrice par les mitoribosomes, à partir de l'ADN mitochondrial. La plupart des complexes enzymatiques (exemple : ATP-synthase) sont formés par la juxtaposition de polypeptides synthétisés dans la mitochondrie et dans le cytosol (le fluide interne de la cellule).
FONCTIONNEMENT : Elle est considérée comme la "centrale énergétique" de la cellule, car c'est là que se déroulent les dernières étapes du cycle respiratoire (en présence d'oxygène, aérobie) qui convertit l'énergie des molécules organiques issues de la digestion (glucose) en énergie directement utilisable par la cellule (ATP). En cas d'absence d'oxygène la cellule utilise la fermentation dans le cytoplasme pour produire l'énergie nécessaire à son fonctionnement, mais c'est un système beaucoup moins efficace, qui dégrade de façon incomplète le substrat. La production d'acide lactique donne lieu, par exemple, à des phénomènes de crampes. L'augmentation de la concentration en ions lactates dans les cellules musculaires est une des raisons de la fatigue après une activité intense. En effet, ces ions lactates changent le pH intracellulaire et modifient de fait les conditions de fonctionnement enzymatiques de la cellule qui ne peut plus travailler correctement. C'est dans la mitochondrie que se déroulent les 2 dernières phases de la respiration cellulaire : le cycle de Krebs (dans la matrice) et la chaîne de transport d'électrons (au niveau de la membrane interne). La première étape, la glycolyse, se déroule dans le cytoplasme cellulaire. Via le cycle de Krebs (donc en condition d'aérobiose), la mitochondrie permet, à partir d'une molécule de glucose, la production de 36 ou 38 molécules d'ATP (cela dépend de la navette utilisée pour transporter le NAD de la glycolyse).
Les mitochondries participent à l'apoptose (mort cellulaire) avec le cytochrome C. De plus, elles ont aussi une fonction de concentration et de stockage des ions calcium, sodium et potassium où ils sont stockés sous forme de granules opaques. On trouve également de l'or, du fer et de l'osmium.
LES MALADIES MITOCHONDRIALES
- Myopathies
- Maladies neurodégénératives
- Ataxie de Friedreich : Maladie touchant la frataxine (Protéine mitochondriale impliquée dans l'imperméabilité des membranes au fer).
WIKIPEDIA > Mai > 2009 |
Quand la Mitochondrie vivait aux Portes d'Asgard |
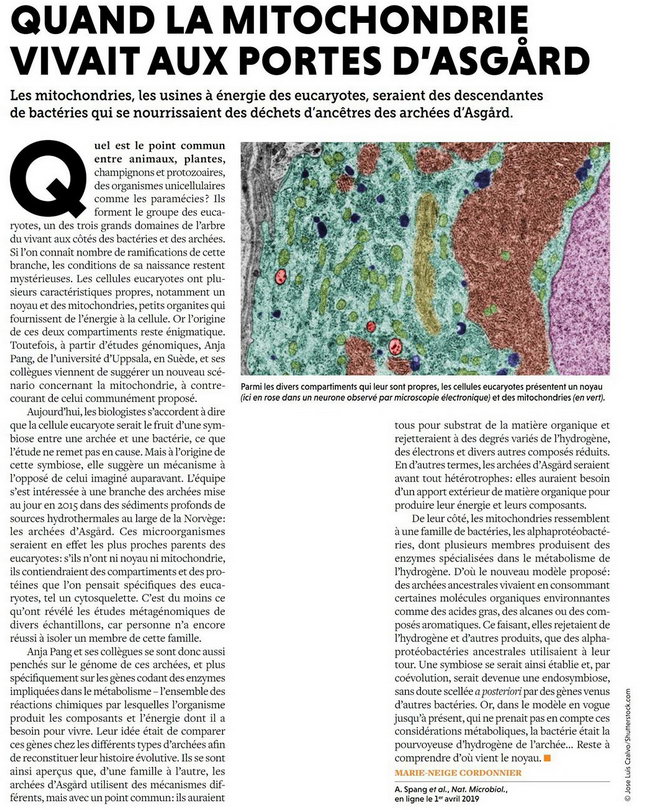
M.N.C. - POUR LA SCIENCE N°500 > Juin > 2019 |
L'Hérédité des Mitochondries Décryptée |

LA RECHERCHE N°515 > Septembre > 2016 |
Vers une Chimie Primitive ? |

L.B.-G. - POUR LA SCIENCE N°485 > Mars > 2018 |
L'ADN des Mitochondries peut aussi venir du Père |

J.-B.V. - SCIENCE & VIE N°1217 > Février > 2019 |